


| name | Amanita magnivelaris |
| name status | nomen acceptum |
| author | Peck |
| english name | "Great Felt Skirt Destroying Angel" |
| images |
 1. Amanita magnivelaris, ca. Ithaca, New York, U.S.A.  2. Amanita magnivelaris, Prov. Quebec, Canada. ") 3. Amanita magnivelaris, Prov. Ontario, Canada. (RET 513-2) |
| intro | The following is based on original description of Peck (1898), a modern description by Lamoureux and Neumann (1990), and the extensive notes on specimens of the species collected near Ithaca by G. F. Atkinson in the early 20th Century (now in the Cornell University Plant Pathology Herbarium, originally misdetermined as A. elliptosperma G. F. Atk.). In addition a collection of R. Singer from Wisconsin (now in the Farlow Herbarium) and several collections sent from Canada by M. Yves Lamoureux were of great value. |
| cap |
The cap is 40 - 127 mm wide, white to cream or yellowish white, viscid when moist, convex at first, later planar and eventually concave in age. It occasionally bears a membranous patch of the volva. Infrequently the entire free limb of the volva is present on the cap in a group of discontinuous patches. |
| gills |
The gills are distinctly attached to the stem or free, close, white, and sometimes have faint pinkish tint. |
| stem |
The stem is 70 - 178 × 7 - 12.5 mm (length includes bulb), white, subcylindric or narrowing upward, and floccose to somewhat scaly. The bulb at the stipe's base is usually greater than 20 mm wide and tapers to a point on its bottom. The very broad ring is felted rather than membranous, unusually thick for species of sect. Phalloideae, white to cream to yellowish cream, floccose on the bottom side, and superior to apical. The volva is limbate and has membranous limbs that are white to grayish and markedly more distant from the stem than in A. bisporigera G. F. Atk. |
| odor/taste | The odor is not reported. This mushroom is deadly POISONOUS. |
| spores | The spores measure (7.2-) 8.2 - 10.8 (-12.5) × (5.2-) 5.8 - 7.8 (-9.2) µm and are broadly ellipsoid to ellipsoid to (occasionally) elongate and amyloid to strongly amyloid. Basida lack clamp connections at their bases as appears to be true throughout sect. Phalloideae. |
| discussion |
The cap of A. magnivelaris does not normally turn yellow in response to KOH solution. However, some specimens have a tendency to turn yellow when bruised or after some period of exposure (possibly due to the "yellowing syndrome"?) and these specimens will give some yellow reaction when tested with KOH. Some authors have called these yellowing specimens "A. decipiens sensu Lamoureux." (See the key to the taxa of sect. Phalloideae in North America, cited below with link.) This species is DEADLY POISONOUS. Amanita magnivelaris was originally described from Long Island, New York. It is known with confidence from southeastern Canada and the adjacent states of the US (Wisconsin to New York). This species is solitary to subgregarious to gregarious in sand or sandy soils (e.g., sandy loam). It is found under oak (Quercus) or in mixed woods of pine (Pinus), Canadian Hemlock (Tsuga canadensis), Beech (Fagus grandifolia) and oak as well as under Birch (Betula). Amanita magnivelaris is not commonly collected and probably should be considered rare. The pointed base of the stem's bulb does not separate this species from members of the A. elliptosperma "group." The spore size and shape is not much help either; however, the very large, felted ring and the floccose stem are very distinctive characters that are useful in the field and in the laboratory. My understanding of the present species owes a great deal to an extensive correspondence over many years with M. Yves Lamoureux (Prov. Quebec, Canada) who persisted in his labors until I finally "got it." Thank you, Yves. The reader may want to examine the recently revised key to the taxa of sect. Phalloideae in North America.—R. E. Tulloss [Note: G. F. Atkinson photos and images from other sources may be located from the Photographic Collections page of the Cornell Plant Pathology Herbarium.] |
| brief editors | RET |
| name | Amanita magnivelaris | ||||||||
| author | Peck. 1897. Rep. (Annual) Regents Univ. State New York New York State Mus. 50: 96. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Great Felt Skirt Destroying Angel" | ||||||||
| synonyms |
≡Amanitina verna f.s. magnivelaris (Peck) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 78, tab. 31 (fig. 4).
≡Amanita verna f.s. magnivelaris (Peck) E.-J. Gilbert [“(Coker) E.-J. Gilbe” (sic)]. 1941. Iconogr. Mycol. (Milan) 27, suppl.: 321. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 166478, 345959 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | NYS (implicit) | ||||||||
| type studies | Jenkins. 1978a. Mycotaxon 7: 31. | ||||||||
| revisions | Lamoureux and Neumann. 1990. Naturaliste Canad. 117(3): 145-159. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of another researcher is based on original research by R. E. Tulloss. | ||||||||
| basidia | from type study of Tulloss: 27 - 47 × 6.2 - 12.8 μm, dominantly 4-sterigmate, rarely 1-sterigmate, sterigmata ??; neither clamps nor proliferating clamps observed. | ||||||||
| universal veil |
from type study of Tulloss: At stipe base, exterior surface layer: 2 -3 hyphal diameters thick (about 18 μm); filamentous undifferentiated hyphae 1.7 - 7.0 μm wide, dominantly longitudinally oriented; vascular hyphae 3.1 - 6.3 μm wide, frequency not recorded. At stipe base, interior: undifferentiated filamentous hyphae 2.8 - 8.4 μm wide, plentiful, loosely interwoven, without dominant orientation; inflated cells terminal and in short chains, with walls apparently up to 1.0 μm thick, subglobose to ellipsoid to broadly clavate to clavate (e.g., 57 - 90 × 27 - 50 μm); vascular hyphae 1.7 - 7.0 μm wide, frequency not recorded. At stipe base, inner surface: not described. from composite of all material revised by Tulloss: At stipe base, exterior surface layer: 2 -3 hyphal diameters thick (about 18 μm in type); filamentous undifferentiated hyphae 1.7 - 11.5 μm wide, branching, at times in fascicles, somewhat to extensively gelatinized, interwoven, dominantly longitudinally oriented in type; vascular hyphae 2.8 - 11.2 μm wide, frequency not recorded. At stipe base, interior: undifferentiated filamentous hyphae 2.5 - 15.2 μm wide, branching, occasionally with slightly thickened walls, with many tip cells slightly inflated, plentiful to dominating, loosely interwoven, without dominant orientation, sometimes coiling; inflated cells terminal and in short chains, with walls apparently up to 1.0 μm thick, subglobose to ellipsoid to broadly clavate to clavate (e.g., 57 - 159 × 20 - 50 μm), sparsely distributed; vascular hyphae 1.7 - 7.0 μm wide, frequency not recorded. At stipe base, inner surface: filamentous undifferentiated hyphae 2.0 - 9.8 μm, branching, somewhat gelatinized; vascular hyphae 1.7 - 8.0 μm, common. | ||||||||
| stipe context |
from type study of Tulloss: longitudinally acrophysalidic, poorly rehydrating; filamentous undifferentiated hyphae 2.1 - 10.5 μm wide; acrophysalides up to 176 × 35 μm (at least); vascular hyphae 3.1 - 9.8 μm wide. composite from all material revised by Tulloss: longitudinally acrophysalidic; filamentous undifferentiated hyphae 2.1 - 10.5 μm wide; acrophysalides up to 182 × 35 μm (at least), some appearing to have walls up to 1.5 μm thick; vascular hyphae 2.4 - 9.8 μm wide. | ||||||||
| partial veil |
from type study of Tulloss: filamentous undifferentiated hyphae 1.5 - 8.0 μm, dominating, interwoven, but with many subradially oriented, branching; inflated cells in clusters, locally common, thin-walled, clavate, terminal or in short chains, e.g. 60 - 139 × 22 - 44 μm; vascular hyphae 1.5 - 6.5 μm, frequency not recorded. composite from all material revised by Tulloss: filamentous undifferentiated hyphae 1.5 - 8.0 μm, dominating, interwoven, but with many subradially oriented, branching; inflated cells in clusters, locally common, clavate, terminal or in short chains, e.g. (49-) 60 - 139 × 15.5 - 44 μm, with walls thin or slightly thickened (up to 0.5 μm thick); vascular hyphae 1.5 - 6.5 μm, frequency not recorded; clamps not observed. | ||||||||
| basidiospores |
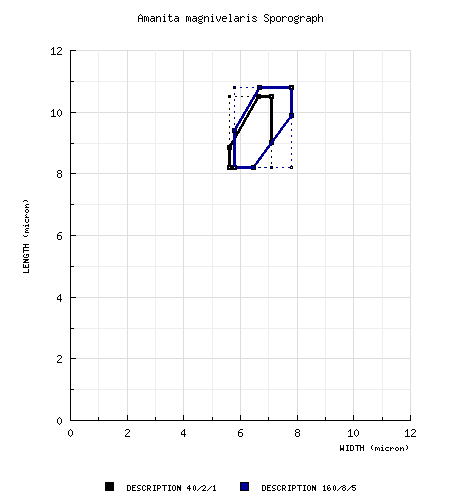
from RET study of type: [40/2/1] (7.8-) 8.2 - 10.5 (-10.6) × (5.3-) 5.6 - 7.1 (-9.2) μm, (L 8.7 - 9.6 μm; L' = 9.1 μm; W = 6.0 - 6.8 μm; W' = 6.4 μm; Q = (1.15-) 1.27 - 1.58 (-1.62); Q = 1.42 - 1.43; Q' = 1.42). composite of spore data from material examined: [160/8/5] (7.2-) 8.2 - 10.8 (-12.5) × (5.2-) 5.8 - 7.8 (-9.2) μm, (L = 8.7 - 9.9 μm; L' = 9.5 μm; W = 6.0 - 7.1 μm; W' = 6.6 μm; Q = (1.15-) 1.27 - 1.62 (-1.72); Q = 1.38 - 1.49; Q' = 1.43), ?? | ||||||||
| ecology | Solitary to subgregarious. Québec: in more or less sandy soil in 200 to 300 year old stand of Beech (Fagus grandifolia). New York: Type probably in deep sandy soil of Pinus rigida-Quercus barrens. Otherwise, in "open woods." Wisconsin: Under Betula. | ||||||||
| material examined |
from type study of Jenkins
(1978a):
U.S.A.:
NEW YORK—Suffolk Co. (Long Isl.) - Port
Jefferson, CANADA: ONTARIO—Port Dover, 2.ix.2012 Eva Skific s.n. [www.mushroomobserver.org #108112 (RET 513-2). QUÉBEC—Laval, 21.viii.1990 Y. Lamoureux s.n. (in herb. Cercle Mycol. Montréal). U. S. A.: NEW YORK—Sussex Co. (Long Isl.) - Port Jefferson, | ||||||||
| discussion |
(t.b.d.) Spore data from the type study of Jenkins (1978a) follows: [-/-/1] (7.3-) 7.9 - 10.2 × (5.8-) 6.3 - 7.9 (-8.4) μm, (Q = 1.13 - 1.46; Q' = 1.27), hyaline, smooth, thin-walled, inamyloid, subglobose to broadly ellipsoid to ellipsoid, adaxially flattened; apiculus sublateral, cylindric to slightly truncate-conic; contents guttulate; color in deposit not recorded. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita magnivelaris |
| name status | nomen acceptum |
| author | Peck |
| english name | "Great Felt Skirt Destroying Angel" |
| images |
1. Amanita magnivelaris, ca. Ithaca, New York, U.S.A. 2. Amanita magnivelaris, Prov. Quebec, Canada. 3. Amanita magnivelaris, Prov. Ontario, Canada. (RET 513-2) |
| photo |
G. F. Atkinson - (1) New York (state), U.S.A., used by
permission of Dr. Kathie Hodge, Director, Plant Pathology Herbarium, Cornell University. Yves Lamoureux - (2) Prov. Québec, Canada. Eva Skific - (3) Prov. Ontario, Canada. (RET 513-2) [Note: The original, unedited version of this image, as well as other images of the same collection, may be found on the mushroomobserver site here.—ed.] |
| drawing | [Note: A superb illustration of the present species can be found on the web site of the Cercle des Mycologues de Montréal (here).] |
| name | Amanita magnivelaris |
| bottom links |
[ Section Phalloideae page. ] [ Amanita Studies home. ] [ Keys & Checklists ] |
| name | Amanita magnivelaris |
| bottom links |
[ Section Phalloideae page. ] [ Amanita Studies home. ] [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.